Canine hyperactivity, impulsivity, and inattention share similar demographic risk factors and behavioural comorbidities with human ADHD
書誌情報Sulkama, S., Puurunen, J., Salonen, M. et al. Canine hyperactivity, impulsivity, and inattention share similar demographic risk factors and behavioural comorbidities with human ADHD. Transl Psychiatry 11, 501 (2021). https://doi.org/10.1038/s41398-021-01626-x
表題の論文を全文日本語訳してみました。翻訳アプリにかけた日本語訳を英文に照らして修正していますが、表記のゆれや訳の間違いがあるかもしれません。正確に内容を知りたい方は、原文をご覧ください。
概要:Abstract
注意欠陥多動性障害(Attention-deficit Hyperactivity Disorder: ADHD)は神経発達障害の一つであり、患者のQOL(Quality of Life)を低下させる。
家庭犬も人間のADHDの構成要素である、高い「多動性・衝動性」「不注意」を自発的に示すことがある。
したがって、犬の多動性・衝動性、不注意に影響を与える人口統計学的、環境的、行動的要因をよりよく理解することは、ヒトと犬の双方に利益をもたらすと考えられる。
我々は、フィンランドの1万1000頭以上の家庭犬から包括的な行動調査データを収集し、多動性・衝動性、不注意のレベルを定量化した。
一般化線形モデル分析を行い、これらの行動特性に関連する因子を同定した。
その結果、多動性・衝動性、不注意が高い犬は、若く、オスで、家で一人で過ごす時間が長い犬において、より多く存在していることが示された。
さらに、これらの特性には遺伝的基盤があることを示唆する、いくつかの犬種の差異が示された。
さらに、多動性・衝動性、不注意は、強迫行為、攻撃性、恐怖心を強く併発することがわかった。
これらの関連性の多くはヒトでも確認されており、ADHDの動物モデルとしての犬の役割が強化されている。
はじめに:Introduction
注意欠陥多動性障害(ADHD)は、遺伝性の高い小児期発症の神経発達障害であり、ヒトにおける世界的有病率は2~7%と推定されている。
ADHDは、注意と報酬処理、抑制的制御、感情制御の異常によって引き起こされる、持続的かつ不適切なレベルの運動過活動、衝動性、不注意の存在によって、特徴づけられる。
ADHDは、多動性・衝動性優位型、不注意優位型、複合型の3種類の提示に分類される。
ADHDはしばしば、学習障害、自閉症スペクトラム、不安障害などのいくつかの併存疾患を伴って成人期まで持続し、適切に診断・治療されないと有害な疾患となる。
人間のADHDの構成要素である衝動性は、運動抑制制御の障害と満足の遅延に耐えられないことによって特徴づけられる。
これは、ヒトやイヌを含む種を超えて観察される正常な性格の連続体の一側面である。
しかし、過剰な衝動性は異常とみなされ、ヒト、げっ歯類、イヌなどのいくつかの種で、攻撃性など他の有害な行動とも関連している。
現在、ADHDの認知度が低いことが、ヒトにおける管理の妨げになっている。
これは、ほとんどがADHDの適切な動物モデルがないことに起因している。
研究によると、12~15%もしくは20%のイヌが、うまれつき高いレベルの多動性・衝動性と不注意を示しており、家庭犬はADHDの動物モデルとして高い可能性を秘めている。
さらに、これらの特性は、イヌとヒトの両方で同じ行動的、生物学的、遺伝的要因によって媒介され、イヌはヒトのADHD治療に用いられる薬剤にも反応する。
さらに、イヌは古典的な動物モデルであるげっ歯類よりも、さらに多くの利点を持っている。
家畜化の過程で、イヌは類似の環境要因にさらされ、収斂的な社会進化を経験した。
また、多くの複雑な社会的認知課題、遺伝学、体の大きさや生理、環境や生活様式の共有においてヒトと同等である。
後者の特徴は、高い遺伝率の推定にもかかわらず、非遺伝的要因もADHDの病因に寄与していることから、イヌをADHDの非常に興味深いモデルとしている。
しかし、どの環境因子が、ADHDの結果にどの程度影響を与えるかはよく分かっていない。
ヒトのADHDに本当に似た研究コホートを得るためには、イヌの客観的で信頼できる表現型が必要である。
これは、飼い主がイヌの行動をよく知っているため、飼い主が記入したアンケートによって達成することができ 、アンケートの信頼性は通常良好である。
Vasらは子どものADHDを測定する調査に基づいて、イヌの多動性、衝動性、注意力を確実に測定できる飼い主記入式の「犬ADHD質問票」を開発した。
この質問票は最近検証され、衝動性と不注意の高さは認知課題の成績低下と関連していた。
本研究では、Vasらによって開発された同じ質問を、犬の包括的行動質問票の一部として利用した。
我々は、フィンランドの1万1000頭以上の家庭犬の研究コホートにおいて、犬の多動性・衝動性と不注意に関連する人口統計学的、環境的、行動的要因を調査することを目的とした。
関連因子の同定は、犬の多動性・衝動性、不注意の異常レベルの予防と管理をより効率的に行うことができ、また、人間のADHD研究にも役立つと思われる。
素材と方法:Material and methods
データ収集
質問票
フィンランドの家庭犬の行動と背景に関する情報を収集するために、飼い主が記入するオンライン行動アンケートを作成した。
質問票には、恐怖、攻撃性、騒音過敏、表面・高所恐怖、多動・衝動性、不注意、分離関連行動、強迫行動の7種類の犬の行動特性に関する質問が含められた。
さらに、アンケートには、犬の生活史に関連する人口統計学的および環境的な質問をカバーする大規模な背景セクションが含まれていた。
質問票の回答は、2015年2月から2018年9月まで収集された。質問票と行動特性分類の詳細は、Salonenらの論文に補足資料として掲載されている。
今回の研究は、Salonenらの論文と同じデータに基づいているが、ここでは大規模な多変量解析を活用した。ここでは、多動性・衝動性と不注意に関連する人口統計学的因子と環境因子を検討した。
多動性/衝動性と不注意
多動性・衝動性と不注意の個人差を測定するために、Vasらが開発し検証した「犬ADHD調査」を用いた。
この調査は、子どものADHDと関連問題の検証された親報告型評価尺度(ADHD RS親バージョン)をベースとしている。
調査には多動性、衝動性、不注意な行動に関する13の記述(項目)(補足表S1)があり、フィンランド語に翻訳されたものである。
犬の飼い主は、その文が自分の犬にとってどれくらいの頻度で当てはまるかを4段階のリッカート尺度(1=決して、2=めったに、3=よく、4=非常によく)で回答するよう求められた。
Vasらの因子構造と同様に、プロマックス回転を用いた主成分分析により、質問文を2つの成分、多動性・衝動性と不注意に分け、それぞれ5文と7文で構成した。
1つの記述(項目11:「急いで反応する可能性が高く、そのために課題を失敗する」)は両方の成分に等しく負荷されるため、分析から除外された。
各犬の多動性・衝動性、不注意の成分スコアを算出し、成分スコアが高いほど多動性・衝動性、不注意のレベルが高いことを示すとした。
記述と構成要素に関する詳細な情報はSalonenらの論文の補足資料に記載されている。質問票のフィンランド語訳は最近Salonenらで検証された。
人口統計学的、行動学的、環境学的変数
統計解析の前に、我々は、行動アンケートから得られたいくつかの人口統計学的変数と環境変数を編集した。
Puurunenら、Hakanenら、Mikkolaらで、すでに記述されているいくつかの新しい変数を作成した。行動アンケートから得られたすべての説明変数は、補足表S2に詳しく説明している。
簡単に説明すると、十分なサンプルサイズを持つ22の犬種と雑種犬を含み、その他の犬種は犬種グループその他でグループ化した。
犬種の平均体高を評価して新たな変数「体格」を作成し、小型犬、中型犬、大型犬に分類した(Supplementary Methods)。
なお、雑種犬については、体格を把握することができないため、体格を解析に含める際には除外した。
犬の居住地の環境土地利用を定量化するために、連続変数「都市環境スコア」を作成し(Supplementary Methods)、都市環境スコアが高いほど、建築環境の割合が高いことを示すようにした。
さらに、「強迫行動」、「攻撃性」、「恐怖心」という3つのカテゴリー的行動変数を作成した(Supplementary Methods)。
3つの特性すべてにおいて、犬を低、中、高の3群に分けた。低グループには強迫的行動、攻撃的行動、恐怖的行動を全く示さない犬を、中グループにはこれらの行動をときどきしか示さない犬を、高グループには少なくとも一つの下位特性で強迫的行動、攻撃的行動、恐怖的行動を定期的に示す犬を入れた。
統計解析
すべての統計解析はRバージョン3.6.2で実施した。
一般化線形モデルは、人口統計学的変数、環境変数、行動変数と多動性・衝動性、不注意との関連を分析するために使用された。
多動性・衝動性と不注意の両モデルには、データに最も適合する対数リンク関数を持つガンマ分布が用いられた。
初期データは264犬種、1万3715頭から構成されていた。
調査した説明変数に欠損や不完全な回答がある個体を除外した結果、多動性・衝動性、不注意ともに6400頭のデータが含まれていた。
解析では、多動性・衝動性、不注意の各成分得点を連続的な回答変数として用いた。
14個の説明変数が先行文献に基づいて分析用に選択された。
人口統計学的説明変数として年齢、性別、不妊手術、犬種、体格が、環境的説明変数として離乳年齢、活動・訓練、日常運動、飼い主の犬の飼育経験、日常一匹で過ごす時間、都市環境スコアが、行動的説明変数として強迫行動、恐怖心、攻撃性が含まれていた。
説明変数として年齢と性を含むモデルから始めて、フォワードステップワイズAIC(赤池情報量規準)モデル選択で最も適合するモデルを探した。
我々の以前の研究では、性別と年齢層は有病率に差があった。
AICモデル選択と最終モデルは補足表S3に示す。
サンプルサイズを最大化するために、最終モデルで選択されなかった説明変数に欠損回答があったすべての犬を含めることによって、モデル選択後に初期データの新しいサブセットを作成した。
その結果、最終的なデータセットは、多動性・衝動性と不注意において、それぞれ1万1539頭と1万1164頭で構成された。
モデルの適合度は慎重に評価された(Supplementary Methods)。
モデルの適合後、推定限界平均をパッケージ ‘emmeans’で計算し、カテゴリ説明変数の調整平均と信頼限界値を得た。Rのパッケージ’effects’はモデル中の他の変数を調整し、連続説明変数の効果を得るために使用された。
多動性・衝動性と不注意スコアに対する説明変数の全体的な効果を得るために、Rのパッケージ’car’ を用いて分散分析(ANOVA)が実施された。
先行研究に基づいて、我々は説明変数のレベル間のいくつかの先験的な対照を形成した。
我々は、若い犬は高齢の犬よりも多動性・衝動性が高く、不注意が強いと仮定した。
また、多動性・衝動性、不注意ともに、頻繁に(少なくとも毎週)活動や訓練に参加している犬と、まったく、あるいはめったに活動や訓練に参加していない犬とは異なるという仮説を立てた。
多動性・衝動性では、大型犬と小型犬では多動性・衝動性が異なるという仮説を立てた。
さらに、高強迫行動群、高攻撃性群、高恐怖性群の犬は、それぞれ低強迫行動群、低攻撃性群、低恐怖性群の犬より多動性・衝動性が高いという仮説を立てた。
Rのパッケージ’emmeans’を使用して、カテゴリ説明変数のレベル間のすべてのペアワイズ比較を調べ、先験的対照を検討した。先験的に選択された対比を除き、偽発見率(FDR)のためにp値を補正した。有意性のカットは、p値<0.05とした。
研究成果:Results
研究コホートと人口統計学
1万1539頭と1万1164頭の犬からなるコホートにおいて、環境、人口統計学的、行動学的要因が多動性・衝動性と不注意に及ぼす影響をそれぞれ検討した。
多動性・衝動性スコアは-1.62から5.23(平均-0.01)、不注意スコアは-1.81から4.81(平均-0.01)とばらつきがあることが判明した。
多動性・衝動性、不注意ともに、51%がメスであった。
犬の年齢は2カ月から17.9歳で、両特徴とも平均年齢は4.7歳であった。
より詳細な人口統計学は補足表S4に示した。
多動性/衝動性に関連する因子
年齢、性別、犬種、体格、毎日の運動量、毎日一匹で過ごす時間、飼い主の飼育経験、強迫行為、攻撃性、恐怖心を含むいくつかの人口統計学的、環境的、行動的変数が、犬の多動性・衝動性と関連していた(表1)。
表1表1:ガンマ分布と対数リンク関数を用いた一般化線形モデルにおける人口統計学的、環境的、行動的変数と多動性/衝動性、不注意スコアとの関連性
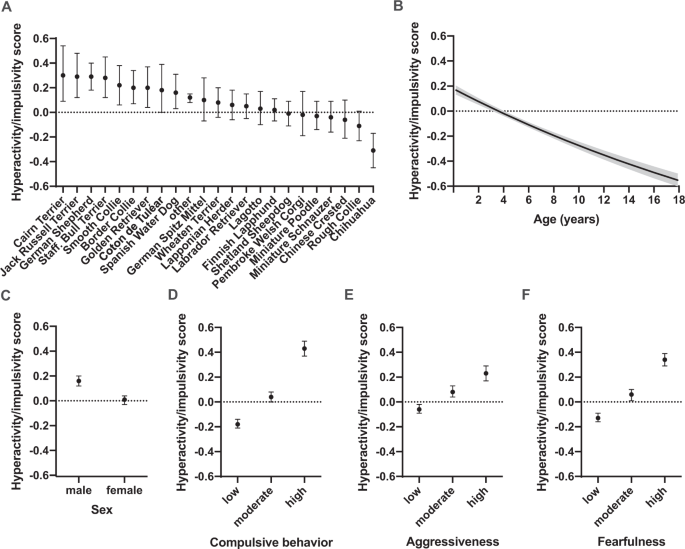
データ中の他のすべての犬種からなるその他のグループを含む23犬種において、多動性・衝動性の犬種間差異を検討した。
その結果、犬種間の多動性・衝動性スコアに有意差が検出された。最もスコアの高い犬種は、
- ケアーン・テリア
- ジャック・ラッセル・テリア
- ジャーマン・シェパード
- スタッフォードシャー・ブル・テリア
であった。最もスコアが低かった犬種は、
- チャイニーズ・クレステッド・ドッグ
- ラフ・コリー
- チワワ
であった(図. 1A)。
チワワとジャーマンシェパードドッグ(z-ratio = -6.07, df = 1, p = 0.0008)、チワワとジャックラッセルテリア(z-ratio = -5.41, df = 1, p = 0.0008 )、チワワとスタッフォードシャー・ブル・テリア(z-ratio = -6.07, df = 0.0008 )のあいだで最大の対差がみられた。
スタッフィー(z比=-5.35、df=1、p=0.0008)およびチワワとその他(z比=-5.22、df=1、p=0.0008)である。すべてのペアワイズ品種差は補足のデータセットに示した。
犬の年齢と性別の両方が多動性・衝動性と関連していた(表1、補足表S5、図1B、C)。
多動性・衝動性得点は、先験的に仮定されたように、若い犬で最も高かった(F = 295.93, df = 1, p < 0.0001)。
雄犬は雌犬より多動性・衝動性スコアが高かった(z-ratio = 8.27, df = 1, p < 0.0001)。
また、体格と多動性・衝動性のあいだには関連があった(補足表S5、補足図S1A)。
中型犬は小型犬(z-ratio = 4.74, df = 1, p = 0.0008)、大型犬(z-ratio = 2.78, df = 1, p = 0.0224)よりも多動・衝動性得点が高く、先験対照で仮定したとおり、小型犬と大型犬のあいだにも有意差が見られた(z-ratio = -2.40, df = 1, p = 0.0166 )。
一日の運動量が少なく、1匹で過ごす時間が長い犬は、多動性・衝動性スコアが高かった(補表S5、補図S1B,C)。
1日の運動時間が1時間未満の犬は、1日の運動時間が3時間以上(z-ratio = 4.75, df = 1, p = 0.0008),2~3 時間(z-ratio = 3.96, df = 1, p = 0.0008),1~2 時間(z-ratio = 2.5, df = 1, p = 0.0416) より多動性・衝動性のスコアが高くなることが示された。
さらに、1~2時間の運動をした犬は、2~3時間(z-ratio = 2.66, df = 1, p = 0.0297)または3時間以上(z-ratio = 3.74, df = 1, p = 0.0014)毎日運動した犬と比較すると、多動・衝動性のスコアが高かった。
毎日8時間以上1匹で過ごす犬は、3時間未満(z-ratio = 3.99, df = 1, p = 0.0008)、3~6時間(z-ratio = 4.83, df = 1, p = 0.0008) または6~8時間(z-ratio = 3.48, df = 1, p = 0.0031) 1匹で過ごす犬より多動・衝動の得点が高いことが示された。
飼い主の飼育経験は、多動性・衝動性スコアの高さと関連していた(補足表S5、補足図S1D)。
飼い主が初めて飼った犬でない場合、飼い主が初めて飼った犬である場合よりも多動性・衝動性スコアが高くなる傾向があった(z-ratio = 4.65, df = 1, p = 0.0001)。
強迫的、攻撃的、恐怖的な犬は多動性・衝動性スコアが高いことが報告された(補表S5、図1D,E,F)。強迫性が高い犬は、低い犬に比べて多動性・衝動性スコアが高かった(z-ratio = 21.50, df = 1, p < 0.0001)。
また、強迫行動のレベルが高い犬は、強迫行動のレベルが中程度の犬と比較して、多動性・衝動性スコアが高く(z-ratio = 13.78, df = 1, p = 0.0008)、強迫行動のレベルが中程度の犬は、強迫行動のレベルが低い犬よりも多動性・衝動性スコアが高くなった(z-ratio = 11.61, df = 1, p = 0.0008 )。
同様に、事前の仮説通り、攻撃性が高い犬は、攻撃性が低い犬と比較して、多動性・衝動性得点が高かった(z-ratio = 10.21, df = 1, p < 0.0001)。
また、攻撃性が高い犬は、攻撃性が中程度の犬よりも多動性・衝動性得点が高く(z-ratio = 4.71, df = 1, p = 0.0008)、攻撃性が中程度の犬は、攻撃性が低い犬よりも多動性・衝動性得点が高い(z-ratio = 6.80, df = 1, p = 0.0008)ことが示された。
さらに、事前仮説どおり、恐怖心が高い犬は、低い犬に比べて多動性・衝動性得点が高かった(z-ratio = 20.01, df = 1, p < 0.0001)。
また、恐怖心のレベルが高い犬は、恐怖心のレベルが中程度の犬と比較して、多動性・衝動性スコアが高く(z-ratio = 10.93, df = 1, p = 0.0008)、恐怖心のレベルが中程度の犬は、恐怖心のレベルが低い犬よりも多動性・衝動性スコアが高かった(z-ratio = 8.77, df = 1, p = 0.0008).
不注意と関連する因子
統計解析により、犬種、年齢、性別、一日に一匹で過ごす時間、活動・訓練、強迫行為、攻撃性、恐怖心など、不注意スコアと関連するいくつかの人口統計学的、環境的、行動的因子が同定された(表1)。
犬種によって不注意スコアに有意差があることが検出された。最もスコアが高かった犬種は、
- ケアーン・テリア
- ゴールデン・レトリーバー
- フィンランド・ラップランド・ドッグ
であった。一方、最もスコアが低かった犬種は、
- スパニッシュ・ウォーター・ドッグ
- ミニチュア・プードル
- ボーダー・コリー
であった(図2A)。
ボーダー・コリーとその他(z-ratio = -8.01, df = 1, p = 0.0005)の間で最も大きなペアワイズ差が認められた。
- ボーダー・コリーとフィンランド・ラップランド・ドッグ(z-ratio = -7.60, df = 1, p = 0.0005)、
- ボーダー・コリーとミックス犬(z-比率=-6.83、df=1、p=0.0005)、および
- ボーダー・コリーとウィーテン・テリア(z-比率 = -6.54, df = 1, p = 0.0005)である。
すべてのペアワイズ品種差は、補足資料に示されている。
犬の年齢は不注意と関連していた(表1、補足図S2A)。不注意のスコアは若い犬で最も高かった(線形効果。F = 8.73, df = 1, p = 0.0031, 二次効果: 線形効果:F = 8.73, df = 1, p = 0.0031, 二次効果:F = 0.56, p = 0.561) 先験的に仮定した通りであった。
また、オスはメスよりも不注意得点が高く、男女間で有意差が見られた(z-ratio = 8.95, df = 1, p < 0.0001; Supplementary Table S6 and Fig. 2B)。
また、一匹で過ごす時間が長く、活動や訓練に参加する頻度が低い犬は、不注意得点が高かった(補足表S6、補足図S2B、C)。
毎日8時間以上一匹で過ごす犬は、一日に3時間未満(z-ratio = 2.99, df = 1, p = 0.0097)、3~6時間(z-ratio = 3.85, df = 1, p = 0.0005) または6~8時間(z-ratio = 2.49, df = 1, p = 0.0327)一匹で過ごす犬より、高い不注意点数をもっていた。
事前に想定したとおり、活動・訓練に参加したことがない/少ない犬と、少なくとも毎週参加する犬では、活動・訓練に参加したことがない/少ない犬の方が不注意得点が高かった(z-ratio = 9.52, df = 1, p < 0.0001 )。
また、ときどき活動に参加する犬は、毎週訓練を受ける犬よりも不注意スコアが高かった(z-ratio = 9.28, df = 1, p = 0.0005)。
強迫的、攻撃的、恐怖的な行動を示す犬も、不注意スコアが高かった(補足表S6、図2C,D,E)。
強迫行為のレベルが高い犬は、強迫行為のレベルが低い(z-ratio = 17.04, df = 1, p = 0.0005)または中程度(z-ratio = 8.63, df = 1, p = 0.0005)の犬と比較すると、不注意スコアがより高かった。
また、中程度の強迫行動を示す犬は、低レベルの強迫行動を示す犬よりも不注意スコアが高かった(z-ratio = 12.17, df = 1, p = 0.0005)。
同様に、攻撃性が高い犬は、攻撃性が低い犬(z-ratio = 7.50, df = 1, p = 0.0005)または中程度(z-ratio = 2.72, df = 1, p = 0.0199)の犬と比較すると、不注意のスコアが高いことが示された。
中程度の攻撃性を示す犬は、低い攻撃性を示す犬よりも不注意得点が高かった(z-ratio = 5.93, df = 1, p = 0.0005)。
また、恐怖心が高い犬は、恐怖心が低い犬(z-ratio = 14.86, df = 1, p = 0.0005)または中程度(z-ratio = 7.90, df = 1, p = 0.0005)の犬と比較すると、不注意スコアが高いことが示された。
恐怖心が中等度の犬は、恐怖心が低度の犬よりも多動性・衝動性スコアが高かった(z-ratio = 6.85, df = 1, p = 0.0005)。
考察:Discussion
我々は1万1000頭以上の犬を対象に、犬の多動性・衝動性と不注意に関する最も広範な調査ベースの研究を行ない、多くの関連する人口統計学的、環境的、行動的要因を特定した。
この研究は、我々の以前の調査と同じデータに基づいてるが、ここでは、複雑な多変量モデルや連続変数としての多動性・衝動性と不注意スコアを含む、より包括的で精度の高いアプローチを使用した。
我々は、以前の研究で観察された行動的な関連性を確認した。
さらに、新しい人口統計学的および環境的な関連性を報告し、表現型の提示における顕著な品種差を観察し、ヒトのADHDで報告されている危険因子および併存疾患との著しい重複を指摘した。
犬の多動性・衝動性、不注意には、年齢、性別、体格など複数の人口統計学的因子が関連していた。
多動性・衝動性、不注意は若い犬で最も一般的であった。両特徴とも年齢とともに減衰するが、多動性・衝動性は不注意よりやや多い。
先行研究でも多動性・衝動性と不注意の行動は、若い犬でずっと多いことが示されている。
しかし、Vasらは衝動性と犬の年齢とのあいだに有意な関連性を見いだせなかった。
本研究では、多動性・衝動性と不注意はメス犬よりもオス犬でより多く見られた。この結果は、男女間に有意差を認めなかった先行研究、あるいはメスの方が多動性・衝動性が高いとした先行研究と矛盾している。
Vasらのみ、オスがメスより衝動的であるとしたが、その差は統計的に有意ではなかった。
ヒトのADHDも同様に小児期に発症し、女児よりも男児に多く見られる。しかし、この性差の理由は不明である。
女子はADHDの優勢な不注意サブタイプに罹患することが多く、ADHDの目に見える兆候を示すことが少ない。
しかしながら、女子はADHDを発現するために、より高い遺伝的危険因子の負担を必要とするかもしれない。
ステロイドホルモンの役割も提案されており、妊娠中に高レベルのテストステロンにさらされると、ドーパミン作動性システムに影響を与え、その結果、男子がADHDになりやすい可能性がある。
これらの出生前ホルモンの影響は、イヌではまだ研究されていない。
これまでの研究では、小型犬はより衝動的であることが示唆されている。
一方、中型犬や大型犬では小型犬よりも高い多動性・衝動性のスコアが観察された。
Wright, Mills & Polluxは、彼らの知見は複数の活動的な小型テリア種から得られたものであると示唆した。
同様に、我々の研究では中型の作業犬種が複数含まれており、より高い衝動性と中型の体格とのあいだに観察された関連を説明できる可能性がある。
これらの犬種は、作業犬としての特性を最大限に発揮するために、活動性、注意力、警戒心を高めるように繁殖されてきた。
体格でモデルを調整すると、本研究で最も多動性・衝動性の高い犬種はあらゆる体格の犬を含み、多くの犬種が互いに有意に異なっていた。
したがって、犬種間の差は体格の差だけでは説明できず、この結果は多動性・衝動性の遺伝的起源をも示している。
20種以上の犬種を対象に多動性・衝動性、不注意を分析したところ、かなりの違いがあることがわかった。
ケアーン・テリア、ジャック・ラッセル・テリア、ジャーマン・シェパード・ドッグ、スタッフォードシャー・ブル・テリア、スムース・コリーの多動性・衝動性のスコアが最も高く、チワワ、ラフ・コリー、チャイニーズ・クレステッド・ドッグ、ミニチュアシュナウザー、プードルは最も低いスコアとなった。
不注意については、ケアーン・テリア、ゴールデン・レトリーバー、フィンランド・ラップドニアン・ドッグ、ミックスブリード、ウィートン・テリアが最も高く、ボーダー・コリー、プードル、スパニッシュ・ウォーター・ドッグ、シェットランド・シープドッグ、ラブラドール・レトリバーのスコアが最も低い結果となった。
さまざまな用途に使用される犬種では異なる行動特性が評価されるため、犬の選択的な繁殖は犬種特有の行動に影響を与えている。
たとえば、ジャーマン・シェパード・ドッグやボーダー・コリーのようないくつかのワーキングドッグ種では、高い活動性、衝動性、注意力が好まれる。これらの犬は通常、高い注意力と反応性により、より良い訓練性と作業能力を持つ。
反対に、チワワ、ラフ・コリー、プードルなど、愛玩犬やショードッグとして人気の犬種においては、こうした特徴は好まれない。
活動的でなく衝動的な犬の方が、あまり活動的ではない生き方をする飼い主が飼うのに楽だからである。
しかし、副次的な効果として、これらの品種では不注意な行動が強まることがある。
興味深いことに、我々の研究では、スムース・コリーは多動性・衝動性のスコアが最も高い犬種の一つであり、一方、ラフ・コリーはスコアが最も低い犬種の一つであった。この2つの犬種は被毛の長さを除けばほとんど同じである。
しかし、現在ではこれらの犬種の使われ方は異なっている。
ラッシーの映画以降、ラフ・コリーはコンパニオン・ドッグとして人気が出たが、スムース・コリーにはこうした人気が出ず、今でも作業犬やドッグスポーツで使われることが多い。
このことも、この近縁種のあいだで観察された多動性・衝動性の違いを説明するかもしれない。
残念ながら、多くの先行研究において個々の犬種ではなく犬種群が用いられているため、我々の結果と先行研究との包括的な比較は困難である。
我々は、多動性・衝動性と不注意に関連するいくつかの環境因子を同定した。
毎日の運動量が少ないこと、活動や訓練への参加が少ないことは、それぞれ多動性・衝動性スコアが高いこと、不注意スコアが高いことと関連していた。
Leyらは、室内で過ごす時間が短い犬は、室内で過ごす時間が長い犬よりも活動的であることを明らかにした。
しかし、彼らは犬の年齢と室内で過ごした時間のあいだに有意な正の相関を確認し、観察された関連性は、通常、高齢の犬は若い犬よりも多くの時間を室内で過ごしたという事実が影響している可能性があることを示唆している。
いくつかの研究により、頻繁に訓練を受けている犬は、それほど頻繁に訓練を受けていない犬よりも不注意になりにくいことが示されている。
運動とエンリッチメントは、犬の種特異的なニーズを満たす方法となり得る。
したがって、犬がより多く運動し、より頻繁に活動や訓練に参加することは、飼い犬のエネルギーやフラストレーションを、飼い主としてのマナーを守りつつ、上手に発散させることができるのである。
したがって、家庭犬において多動性・衝動性、不注意が高い原因として、エネルギーを発散させたり、活動量を減らしたりする機会が限られているためであるケースも考えられる。
しかし、飼い主が不注意な犬とのトレーニングや活動への参加に消極的なことも多く、その理由は、犬の集中力が低いため、不快で満足のいく成果を得ることに苦労するからである。
ヒトのADHDに対する運動の効果を調査した研究は少なく、サンプルサイズも小さいが、ADHDの子どもにおいては、いくつかのメタアナリシスにより、身体運動が多動性・衝動性、不注意をある程度緩和することが示されている。
私たちは、毎日一匹で過ごす時間が長い犬ほど、一匹で過ごす時間が短い犬よりも、多動性・衝動性、不注意のスコアが高いという新しい知見を観察した。
犬は社会的な動物であるため、長時間一人にされるとストレスや欲求不満になることがある。
このストレスやフラストレーションが、多動性・衝動性、不注意な行動として噴出することがある。
一般に、犬は飼い主と離れているあいだ、落ち着いて休んでいるものである。
分離期間が長くなると、飼い主が戻ってきたときに、より元気な行動や運動量が増えることがあるが、これは分離期間が長くなることで犬の社会的孤立感が増すことを反映している可能性がある。
Rehn & Keelingは、分離時間が長くなると犬はより強い挨拶行動をするようになり、身体活動や注意行動の頻度が高くなる傾向があり、放置時間の影響が確認されたとしている。
しかし、一匹で過ごす時間が長い犬は、それ以外でも飼い主からの注意や運動が少ない可能性もある。
一人で過ごす時間が長いほど多動・衝動的な行動や不注意な行動が多くなることとの関係については、それ以上の結論は出ず、因果関係は推測するしかない。
新たな知見として、飼い主の2頭目以降の犬で多動性・衝動性が高いことを発見した。
このような関連は過去に報告されていないため、これらの要因の関係については推測するしかない。愛玩犬のような活動性の低い犬種のなかから、飼いやすい個体を最初の犬として選ぼうとするのかもしれない。
そして、1頭目の飼育で犬の扱い方の経験を積むことで、より活動的で難しい犬種、たとえば牧畜犬種を次の犬として選びやすくなるのではないだろうか。
また、初めての犬でドッグスポーツや趣味に挑戦し、そのスポーツに夢中になれば、そのスポーツでより活躍できるよう、より活動的で運動量の多い犬種を後住犬に選ぶこともあるだろう。
また、1頭以上の犬を飼っている飼い主は経験が豊富で、犬の多動性・衝動性と不注意などの異なる行動をよりよく認識できているから、という可能性がある。
興味深いことに、強迫行為、攻撃性、恐怖心が強い犬では、多動性・衝動性、不注意のスコアがともに有意に高くなった。
犬において衝動性は、長いあいだ攻撃性と関連していると考えられてきた。
最近のある研究では、強迫行為と多動性、そして恐怖心と多動性のあいだに関連があることが発見された。
さらに、Wrightらは、他の行動上の問題があると報告された犬では衝動性がより一般的であると記述している。
しかし、これらの行動上の問題が何であるかは公表されていない。
しかし、犬では不注意はほとんど研究されておらず、観察されたこれらの併存性(不注意と強迫行動、攻撃性、恐怖心のあいだ)は前、同じデータセットを拡張して行った我々の調査においてのみ記述されていた。
本研究では、これらの共存関係は、いくつかの人口統計学的変数と環境変数を同じ多変量モデルに含めた後でも持続した。
我々の結果と平行して、ADHDはヒトでは自閉症スペクトラム、学習障害、不安・気分障害など、いくつかの行動的併存疾患を持つことが知られている。
ADHDの主要な構成要素である衝動性は、攻撃的な行動とも関連する。
これは、しばしば衝動的攻撃性または反応性攻撃性に分類されることがある。
さらに、強迫性障害(OCD)はヒトにおいてしばしばADHDと併存し、両疾患は抑制的制御の障害と実行機能の欠損によって特徴づけられる。
多動性・衝動性と攻撃性、恐怖心、強迫行為との併存関係は、これらの特徴に関与する共通の基礎的な神経生物学的経路と脳構造を参照している可能性がある。
ADHDと衝動的行動は、前頭葉回路における欠陥と、例えば前頭前野(PCF)、前帯状皮質(ACC)、線条体における異常なレベルの活性化と関連している。
同様に、強迫性障害も前頭葉回路活動の異常によって特徴付けられ、同様に線条体、PFC、ACCが関与するADHDとOCDに加えて、攻撃性も脳の報酬系が関与し、攻撃性に関わる神経経路も同様にPFCと線条体に接続している。最後に、恐怖と不安もPFCとACCにおける活動と関連している。
我々の結果は、異なる犬種がADHDの異なる提示をモデル化するのに有用であることを示している。
ケアーン・テリアは、多動性・衝動性と不注意形質の両方の平均スコアが高く、強迫行為と攻撃性が共存しているので、ADHDのモデルとして適している可能性がある。
一方、ラブラドール・レトリーバーは両特徴の平均点が低く、また、恐怖心、攻撃性、強迫行為などの共存行動を示すことは非常にまれであった。
スパニッシュ・ウォーター・ドッグは不注意の平均点が低いが、多動性・衝動性の平均点は高く、恐怖心などの併存行動もある。
一方、チャイニーズ・クレステッド・ドッグは、不注意の平均点は高いが、多動性・衝動性の平均点は低く、恐怖心、攻撃性、強迫行動も高いレベルであった。
さいごに、多動性・衝動性と不注意に関連する遺伝的・生物学的要因を明らかにするためには、品種内研究が有効であると考えられる。
本研究には限界がある。我々の分析はアンケートに基づいており、飼い主の研究への参加は任意であった。
アンケートは、その信頼性が行動科学において有用であり、アンケート回答が動物の行動と強く結びついていることから、データ収集の効果的な方法となりうる。
しかし、アンケートは主観的になりやすい。
我々のデータは自分で選んだ便宜的なサンプルであり、フィンランドの犬全体の人口を代表していない可能性がある。
データの欠落のために、いくつかの犬は分析から除外されたので、将来の研究は、より完全なデータを収集することを目的とする必要がある。
最後に、調査した犬種のなかには、ワーキングラインとショーラインの両方が含まれているものがあり、犬種内のラインを分けることができなかった。
今後の研究では、ショーラインとワーキングラインのあいだで起こりうる行動のちがいを評価するために、その犬のラインに関する情報を収集することを目指す。
結論として、犬の多動性・衝動性と不注意は、いくつかの人口統計学的、環境的、行動学的因子と関連していることが示された。
また、これらの形質には強い遺伝的基盤があることが示唆された。
さらに、我々の結果は、犬がADHDの適切な、新進気鋭の動物モデルであることを補強するものである。
多動性・衝動性と不注意は、若いオスの犬でより多くみられ、年齢と性別の影響は、人間のADHDでも十分に立証されている。
さらに、イヌの多動性・衝動性、不注意と、ヒトのADHDにおける類似した行動の併存は、両種においてこれらの形質の背後にある共通の神経生物学的経路という仮説を強めるものである。
さらに、犬と人間の遺伝学、生理学、生活環境が類似していることから、犬は、たとえばげっ歯類よりもADHDに対する本質的なモデルであると言える。
したがって、犬の多動性・衝動性と不注意に影響を与える要因を理解することは、犬におけるこれらの特性の認識と管理だけでなく、人間のADHD研究にも有益であると考えられる。
References
Sayal K, Prasad V, Daley D, Ford T, Coghill D. ADHD in children and young people: prevalence, care pathways, and service provision. Lancet Psychiatry. 2018;5:175–86.
Article PubMed Google ScholarFaraone SV, Perlis RH, Doyle AE, Smoller JW, Goralnick JJ, Holmgren MA, et al. Molecular genetics of attention-deficit/hyperactivity disorder. Biol Psychiatry. 2005;57:1313–23.
Article CAS PubMed Google ScholarAmerican Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®). 5th ed. American Psychiatric Publishing; 2013.
Gallo EF, Posner J. Moving towards causality in attention-deficit hyperactivity disorder: overview of neural and genetic mechanisms. Lancet Psychiatry. 2016;3:555–67.
Article PubMed PubMed Central Google ScholarShaw P, Stringaris A, Nigg J, Leibenluft E. Emotion dysregulation in attention deficit hyperactivity disorder. Am J Psychiatry. 2014;171:276–93.
Article PubMed PubMed Central Google ScholarDougherty DM, Bjork JM, Harper RA, Marsh DM, Moeller FG, Mathias CW, et al. Behavioral impulsivity paradigms: a comparison in hospitalized adolescents with disruptive behavior disorders. J Child Psychol Psychiatry Allied Discip. 2003;44:1145–57.
Article Google ScholarRiemer S, Mills D. S. & Wright H. Impulsive for life? The nature of long-term impulsivity in domestic dogs. Anim. Cogn. 2014.
https://doi.org/10.1007/s10071-013-0701-4.Cherek DR, Moeller FG, Dougherty DM, Rhoades H. Studies of violent and nonviolent male parolees: II. Laboratory and psychometric measurements of impulsivity. Biol Psychiatry. 1997;41:523–9.
Article CAS PubMed Google ScholarWinstanley CA, Eagle DM, Robbins TW. Behavioral models of impulsivity in relation to ADHD: translation between clinical and preclinical studies. Clin Psychol Rev. 2006;26:379–95.
Article PubMed PubMed Central Google ScholarOdum AL. Delay discounting: trait variable? Behav Process. 2011;87:1–9.
Article Google ScholarVan den Bergh F, Spronk M, Ferreira L, Bloemarts E, Groenink L, Olivier B. et al. Relationship of delay aversion and response inhibition to extinction learning, aggression, and sexual behaviour. Behav Brain Res. 2006;175:75–81.
Article PubMed Google ScholarCervantes MC, Delville Y. Serotonin 5-HT 1A and 5-HT 3 receptors in an impulsive-aggressive phenotype. Behav Neurosci. 2009;123:589–8.
Article CAS PubMed Google ScholarReisner IR, Mann JJ, Stanley M, Huang YY, Houpt KA. Comparison of cerebrospinal fluid monoamine metabolite levels in dominant-aggressive and non-aggressive dogs. Brain Res. 1996;714:57–64.
Article CAS PubMed Google ScholarPeremans K, Audenaert K, Coopman F, Blanckaert P, Jacobs F, Otteet A, et al. Estimates of regional cerebral blood flow and 5-HT2A receptor density in impulsive, aggressive dogs with 99mTc-ECD and 123I-5-I-R91150. Eur J Nucl Med Mol Imaging. 2003;30:1538–46.
Article CAS PubMed Google ScholarAmat M, Le Brech S, Camps T, Torrente C, Mariotti VM, Ruiz JL, et al. Differences in serotonin serum concentration between aggressive English cocker spaniels and aggressive dogs of other breeds. J Vet Behav Clin Appl Res. 2013;8:19–25.
Article Google ScholarBrady K, Hewison L, Wright H, Zulch H, Cracknell N, Millset D. A spatial discounting test to assess impulsivity in dogs. Appl Anim Behav Sci. 2018;202:77–84.
Article Google ScholarWright HF, Mills DS, Pollux PMJ. Behavioural and physiological correlates of impulsivity in the domestic dog (Canis familiaris). Physiol Behav. 2012;105:676–82.
Article CAS PubMed Google ScholarMajdak P, Ossyra JR, Ossyra JM, Cobert AJ, Hofmann GC, Tse S, et al. A new mouse model of ADHD for medication development. Sci Rep. 2016;6:39472
Article CAS PubMed PubMed Central Google ScholarDinwoodie IR, Dwyer B, Zottola V, Gleason D, Dodman NH. Demographics and comorbidity of behavior problems in dogs. J Vet Behav. 2019;32:62–71.
Article Google ScholarSalonen M, Sulkama S, Mikkola S, Puurunen J, Hakanen E, Tiiraet K, et al. Prevalence, comorbidity, and breed differences in canine anxiety in 13,700 Finnish pet dogs. Sci Rep. 2020;10:2962
Article CAS PubMed PubMed Central Google ScholarVas J, Topál J, Péch É, Miklósi Á. Measuring attention deficit and activity in dogs: a new application and validation of a human ADHD questionnaire. Appl Anim Behav Sci. 2007;103:105–17.
Article Google ScholarLit L, Schweitzer JB, Iosif AM, Oberbauer AM. Owner reports of attention, activity, and impulsivity in dogs: a replication study. Behav Brain Funct. 2010;6:1.
Article PubMed PubMed Central Google ScholarPiotti P, Satchell LP, Lockhart TS. Impulsivity and behaviour problems in dogs: a reinforcement sensitivity theory perspective. Behav Process. 2018;151:104–10.
Article Google ScholarHejjas K, Kubinyi E, Ronai Z, Szekely A, Vas J, Mikl¢si A, et al. Molecular and behavioral analysis of the intron 2 repeat polymorphism in the canine dopamine D4 receptor gene. Genes Brain Behav. 2009;8:330–6.
Article CAS PubMed Google ScholarPuurunen J, Sulkama S, Tiira K, Araujo C, Lehtonen M, Hanhineva K, et al. A non-targeted metabolite profiling pilot study suggests that tryptophan and lipid metabolisms are linked with ADHD-like behaviours in dogs. Behav Brain Funct. 2016;12:27
Article PubMed PubMed Central CAS Google ScholarWan M, Hejjas K, Ronai Z, Elek Z, Sasvari-Szekely M, Champagne FA, et al. DRD4 and TH gene polymorphisms are associated with activity, impulsivity and inattention in Siberian Husky dogs. Anim Genet. 2013;44:717–27.
Article CAS PubMed Google ScholarLuescher UA. Hyperkinesis in dogs: six case reports. Can Vet J. 1993;34:368–70.
CAS PubMed PubMed Central Google ScholarLindsay S. R. Handbook of applied dog behavior and training: etiology and assessment of behavior problems. Iowa: Iowa State University Press; 2001.
Topál J, Miklósi Á, Gácsi M, Dóka A, Pongrácz P, Kubinyi E, et al. The dog as a model for understanding human social behavior. Adv Study Behav. 2009;39:71–116.
Article Google ScholarKaminski J, Call J, Fischer J. Word learning in a domestic dog: Evidence for ‘fast mapping’. Science. 2004;304:1682–3.
Article CAS PubMed Google ScholarLindblad-Toh K, Wade CM, Mikkelsen TS, Karlsson EK, Jaffe DB, Kamal M. et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature. 2005;438:803–19.
Article CAS PubMed Google ScholarThapar A, Cooper M, Eyre O, Langley K. What have we learnt about the causes of ADHD? J Child Psychol Psychiatry Allied Discip. 2013;54:3–16.
Article Google ScholarBennett SL, Litster A, Weng HY, Walker SL, Luescher AU. Investigating behavior assessment instruments to predict aggression in dogs. Appl Anim Behav Sci. 2012;141:139–48.
Article Google ScholarTiira K, Lohi H. Reliability and validity of a questionnaire survey in canine anxiety research. Appl Anim Behav Sci. 2014;155:82–92.
Article Google ScholarDuffy DL, Serpell JA. Predictive validity of a method for evaluating temperament in young guide and service dogs. Appl Anim Behav Sci. 2012;138:99–109.
Article Google ScholarDuPaul GJ, Power TJ, Anastopoulos AD, Reid R. ADHD rating scale—IV: checklists, norms, and clinical interpretation. New York: Guilford Press; 1998.
Bunford N, Csibra B, Peták C, Ferdinandy B, Miklósi Á, Gácsi M. Associations among behavioral inhibition and owner-rated attention, hyperactivity/impulsivity, and personality in the domestic dog (Canis familiaris). J Comp Psychol. 2019;133:233–43.
Article PubMed Google ScholarSalonen M, Mikkola S, Hakanen E, Sulkama S, Puurunen J, Lohi H. Reliability and validity of a dog personality and unwanted behavior survey. Animals. 2021;11:1–16.
Article Google ScholarPuurunen J, Hakanen E, Salonen MK, Mikkola S, Sulkama S, Araujo C, et al. Inadequate socialisation, inactivity, and urban living environment are associated with social fearfulness in pet dogs. Sci Rep. 2020;10:3527
Article CAS PubMed PubMed Central Google ScholarHakanen E, Mikkola S, Salonen M, Puurunen J, Sulkama S, Araujo C, et al. Active and social life is associated with lower non-social fearfulness in pet dogs. Sci Rep. 2020;10:1–13.
Article CAS Google ScholarMikkola S, Salonen M, Puurunen J, Hakanen E, Sulkama S, Araujo C, et al. Aggressive behaviour is affected by demographic, environmental and behavioural factors in purebred dogs. Sci Rep. 2021;11:9433
Article CAS PubMed PubMed Central Google ScholarR Core Team. R: A language and environment for statistical computing. (2019).
Lenth, R. emmeans: estimated marginal means, aka least-squares means. 2019. https://cran.r-project.org/package=emmeans. Accessed 22nd May 2019.
Fox J. Effect displays in R for generalised linear models. J Stat Softw. 2003;8:1–27.
Article Google ScholarFox J, Weisberg S. An R companion to applied regression. Thousand Oaks, CA: Sage Publications; 2011.
Kubinyi E, Turcsán B, Miklósi Á. Dog and owner demographic characteristics and dog personality trait associations. Behav Process. 2009;81:392–401.
Article Google ScholarLey JM, Bennett PC, Coleman GJ. A refinement and validation of the Monash Canine Personality Questionnaire (MCPQ). Appl Anim Behav Sci. 2009;116:220–7.
Article Google ScholarWright HF, Mills DS, Pollux PMJ. Development and validation of a psychometric tool for assessing impulsivity in the domestic dog (Canis familiaris). Int J Comp Psychol. 2011;24:210–25.
Article Google ScholarKubinyi E, Vas J, Hejjas K, Ronai Z, Br£der I, Turcs n B, et al. Polymorphism in the tyrosine hydroxylase (TH) gene is associated with activity-impulsivity in German Shepherd Dogs. PLoS ONE. 2012;7:e30271
Article CAS PubMed PubMed Central Google ScholarTonoike A, Nagasawa M, Mogi K, Serpell JA, Ohtsuki H, Kikusui T. Comparison of owner-reported behavioral characteristics among genetically clustered breeds of dog (Canis familiaris). Sci Rep. 2015;5:17710
Article CAS PubMed PubMed Central Google ScholarPolanczyk G, de Lima MS, Horta BL, Biederman J, Rohde LA. The worldwide prevalence of ADHD: a systematic review and metaregression analysis. Am J Psychiatry. 2007;164:942–8.
Article PubMed Google ScholarBiederman J, Petty CR, Woodworth KY, Lomedico A, Hyder LL, Faraone SV. Adult outcome of attention-deficit/hyperactivity disorder: a controlled 16-year follow-up study. J Clin Psychiatry. 2012;73:941–50.
Article PubMed Google ScholarNewcorn JH, Halperin JM, Jensen PS, Abikoff HB, Arnold LE, Cantwell DP. et al. Symptom profiles in children with ADHD: effects of comorbidity and gender. J Am Acad Child Adolesc Psychiatry. 2001;40:137–46.
Article CAS PubMed Google ScholarMartin J, Taylor MJ, Rydell M, Riglin L, Eyre O, Lu Y, et al. Sex-s pecific manifestation of genetic risk for attention deficit hyperactivity disorder in the general population. J Child Psychol Psychiatry Allied Discip. 2018;59:908–16.
Martel MM, Klump K, Nigg JT, Breedlove SM, Sisk CL. Potential hormonal mechanisms of attention-deficit/hyperactivity disorder and major depressive disorder: a new perspective. Hormones Behav. 2009;55:465–79.
Article CAS Google ScholarWaddell J, McCarthy MM. Sexual differentiation of the brain and ADHD: what is a sex difference in prevalence telling us? Curr Top Behav Neurosci. 2012;9:341–60.
Article PubMed PubMed Central Google ScholarSvartberg K. Breed-typical behaviour in dogs – Historical remnants or recent constructs? Appl Anim Behav Sci. 2006;96:293–313.
Article Google ScholarVaysse A, Ratnakumar A, Derrien T, Axelsson E, Rosengren Pielberg G, Sigurdsson S, et al. Identification of genomic regions associated with phenotypic variation between dog breeds using selection mapping. PLoS Genet. 2011;7:10.
Gerencsér L, Bunford N, Moesta A, Miklósi Á. Development and validation of the Canine reward responsiveness scale -Examining individual differences in reward responsiveness of the domestic dog. Sci Rep. 2018;8:4421.
Article PubMed PubMed Central CAS Google ScholarGhirlanda S, Acerbi A, Herzog H. Dog movie stars and dog breed popularity: a case study in media influence on choice. PLoS ONE. 2014;9:9.
Article CAS Google ScholarCerrillo-Urbina AJ, García-Hermoso A, Sánchez-López M, Pardo-Guijarro MJ, Santos Gómez JL, Martínez-Vizcaíno V. The effects of physical exercise in children with attention deficit hyperactivity disorder: a systematic review and meta-analysis of randomized control trials. Child Care Health Dev. 2015;41:779–88.
Zang Y. Impact of physical exercise on children with attention deficit hyperactivity disorders: evidence through a meta-analysis. Medicine. 2019;98:e17980.
Article PubMed PubMed Central Google ScholarRehn T, Keeling LJ. The effect of time left alone at home on dog welfare. Appl Anim Behav Sci. 2011;129:129–135.
Article Google ScholarKooij JJS, Bijlenga D, Salerno L, Jaeschke R, Bitter I, Balázs J, et al. Updated European Consensus Statement on diagnosis and treatment of adult ADHD. Eur Psychiatry. 2019;56:14–34.
Rosell DR, Siever LJ. The neurobiology of aggression and violence. CNS Spectr. 2015;20:254–79.
Article PubMed Google ScholarHaller J. The neurobiology of abnormal manifestations of aggression-A review of hypothalamic mechanisms in cats, rodents, and humans. Brain Res Bull. 2013;93:97–109.
Article CAS PubMed Google ScholarWaltes R, Chiocchetti AG, Freitag CM. The neurobiological basis of human aggression: a review on genetic and epigenetic mechanisms. Am J Med Genet Part B Neuropsychiatr Genet. 2016;171:650–75.
Article Google ScholarBrem S, Grünblatt E, Drechsler R, Riederer P, Walitza S. The neurobiological link between OCD and ADHD. Atten Deficit Hyperact Disord. 2014;6:175–202.
Norman LJ, Carlisi C, Lukito S, Hart H, Mataix-Cols D, Radua J, et al. Structural and functional brain abnormalities in attention-deficit/hyperactivity disorder and obsessive-compulsive disorder: a comparative meta-analysis. JAMA Psychiatry. 2016;73:815–25.
Tripp G, Wickens JR. Neurobiology of ADHD. Neuropharmacology. 2009;57:579–89.
Article CAS PubMed Google ScholarNakao T, Okada K, Kanba S. Neurobiological model of obsessive-compulsive disorder: evidence from recent neuropsychological and neuroimaging findings. Psychiatry Clin Neurosci. 2014;68:587–605.
Article PubMed Google ScholarMilad MR, Rauch SL. Obsessive-compulsive disorder: beyond segregated cortico-striatal pathways. Trends Cogn Sci. 2012;16:43–51.
Article PubMed Google ScholarSharma A, Couture J. A review of the pathophysiology, etiology, and treatment of attention-deficit hyperactivity disorder (ADHD). Ann Pharmacother. 2014;48:209–25.
Article PubMed Google ScholarLangen M, Durston S, Kas MJH, van Engeland H, Staal WG. The neurobiology of repetitive behavior:…and men. Neurosci Biobehav Rev. 2011;35:356–65.
Article PubMed Google ScholarDel Casale A, Kotzalidis GD, Rapinesi C, Serata D, Ambrosi E, Simonetti A, et al. Functional neuroimaging in obsessive-compulsive disorder. Neuropsychobiology. 2011;64:61–85.
DeYoung CG. Personality neuroscience and the biology of traits. Soc Personal Psychol Compass. 2010;4:1165–80.
Article Google ScholarTovote P, Fadok JP, Lüthi A. Neuronal circuits for fear and anxiety. Nat Rev Neurosci. 2015;16:317–31.
Article CAS PubMed Google Scholar
Keywords:
ADHD, Psychology
{kind=link}